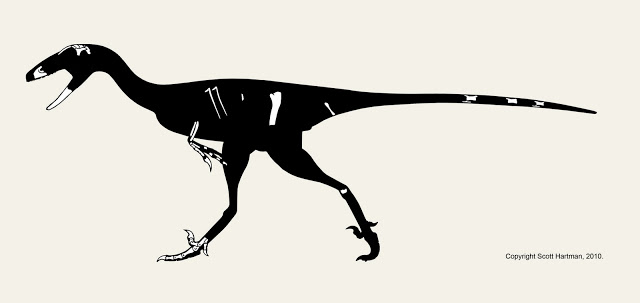
If you watched Episode II of Dinosaur Revolution, you may have laughed at this Ornitholetes, who I'll refer to as Ichabod. This may seem like an odd scene to pick for a scientific discussion, but I think it actually has something useful to teach. Also, I'm partially responsible. I should be clear, the story idea was not mine (that's above my pay grade), but it's something that was run by me, and I did not try to shoot it down (and still wouldn't).
Why? I think it's reasonable.
No wait, let me explain: I'll grant you that there isn't much in the professional literature on the subject on the subject of "headless running" in animals, but from some criticisms I've read I think people maybe thinking about this the wrong way (i.e., wondering about the distribution of "headless-running" in birds like the behavior it's an advanced condition).
Vertebrates as a group have one of the more centralized nervous systems among animals (with some arthropods and especially some cephalopods as the other contestants in the "flexibility over redundancy sweepstakes"), but tetrapod nervous system evolution in general is a story of progressive centralization that (so far) culminates in mammals. Even in humans, with our gobs of ridiculously calorie-hungry centralized gray matter, we still have autonomous reactions that don't require the brain to be involved (as anyone knows who's burned themselves and jerked their hand away before they felt the pain). That said, we have gone a long way down the path of nervous system centralization, and if you cut a mammal's head off you may get some twitching but it won't run around; our limbs literally cannot coordinate themselves without the brain's involvement (although morbidly it does appear that the head itself retains some coordination afterwards, if medieval reports are true that heads react for up to a 15 seconds after a beheading).
This degree of centralization is the derived condition, not the primitive one. So it seems unlikely that chickens are special here, except in as much as they more frequently get clean beheadings in the presence of human observers than most other birds (a quick Google search shows that turkey's exhibit this as well). This should be true of lizards, crocs, etc too (diapsids as a whole). So with enough experimental trials I'd fully expect an Ornnitholestes to do a good headless chicken impression. Now, I'll grant you that this would require a pretty clean bite on the allosaur's part, and the odds of observing it in the wild would not be very high. But the sequence was devised as be a surprising bit of humor in a scenario that was possible, not probable.
Given those parameters it seems reasonable enough to me.



.jpg)




























{kind=link}