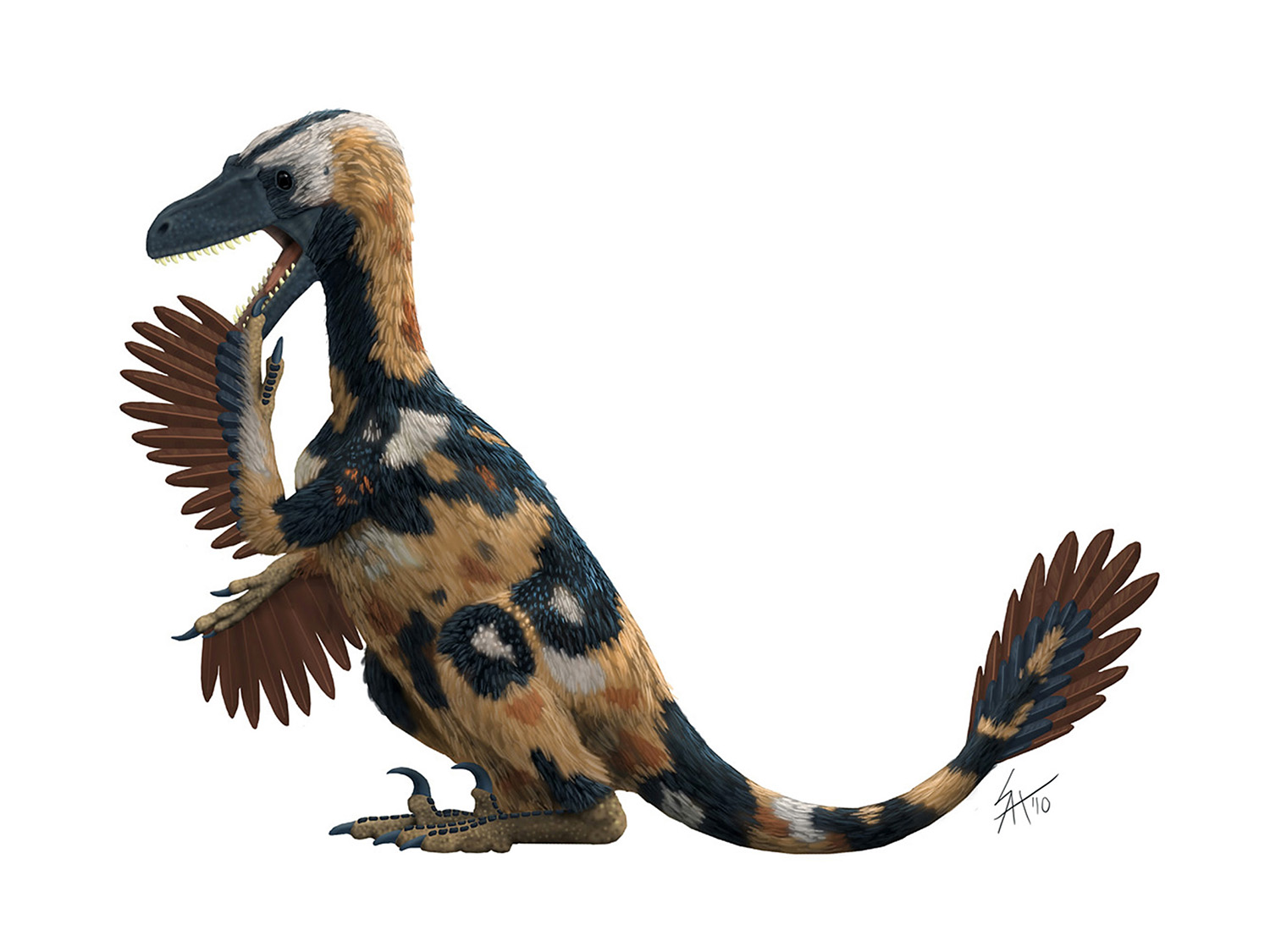Falcarius: bizarre sickle-cutter
/
The truly strange looking animal above is Falcarius utahensis. It's an early, omnivorous member of the theropod clade known as therizinosaurs. Not only does it look weird, it's also a bit different from other skeletals you may have seen on the web. Join me after the break for a bit of a discussion about Falcarius, and the challenges I faced with this reconstruction.
I should warn you, this won't be a tutorial on how I make my skeletal reconstructions. That would certainly be a fun series, but it would require quite a lot of time to do properly, so for now it'll have to wait. But there are still several points worthy of discussion.
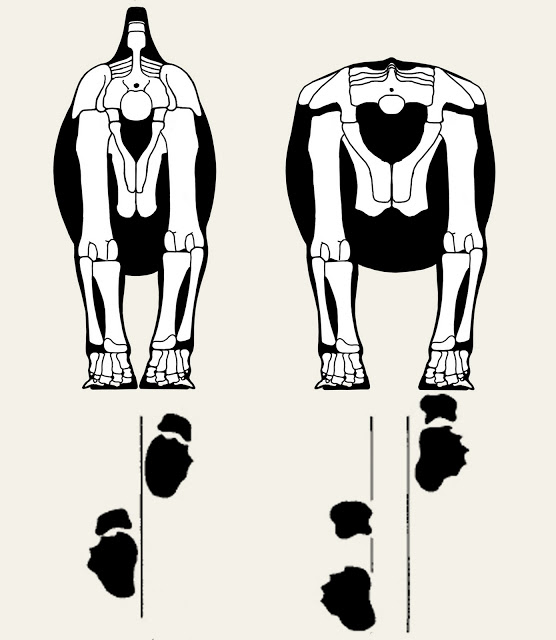
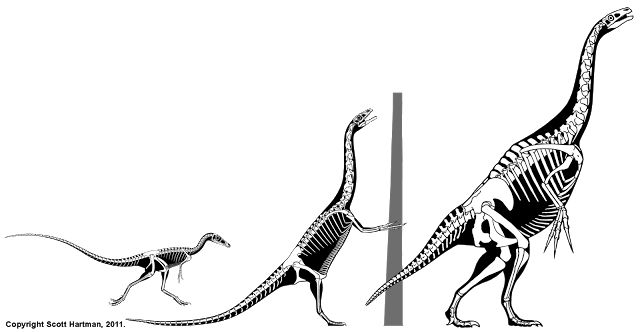
First off, the animal was discovered in a bone bed of disarticulated individuals. The good news is that most of the individual elements are known, but the down side is the bones aren't all from the same sized animals. That means that cross-scaling is needed to restore the skeleton, but even that presents a challenge; the usual method of cross-scaling involves double-checking the results against the proportions of close relatives. Alas, in this case the fossil record for the base of the therizinosaur family tree isn't well known, and what is known makes it clear that Falcarius has very different proportions than it's closest known relative: Beipiaosaurus.

(Image copyright Greg Paul)
When the original description of Falcarius was published in 2005, it came with the skeletal drawing at left. Obviously I don't agree with those proportions now, but at the time it had been done when fewer bones had been excavated, prepared, and described in detail, so Greg Paul had to try and scale them based on a smaller amount of material to compare with.
In fact, given the difficulty of restoring the proportions I intentionally avoided doing a Falcarius skeletal reconstruction for several years. I might have avoided it all together, but towards the end of my tenure at the WDC we mounted a cast of Falcarius that Gaston Design produced. Working on that skeleton I was able to not only measure and photograph all of the elements, but spend time looking at how the individual elements were matched up. Some parts of the cast's vertebral column are from different sized individuals (an unavoidable consequence of trying to piece together a skeleton from several different individuals). In other cases, vertebrae I had assumed to be from different sized animals were in fact crushed.
.jpg)
In addition to the hands-on data, Lindsay Zanno had been hard at work publishing more detailed information on Falcarius (this is actually notable, as not all researchers are as timely with getting more detailed descriptions of a new animal into print).
As the information piled up I felt that a skeletal was possible to be done. I still didn't tackle it though, as there were plenty of "low-hanging fruit" skeletals that could be done from less-challenging animals.
As luck would have it, I ended up being asked to produce a skeletal of Falcarius for a display in the new Utah Museum of Natural History building (side note: the new UMNH building just opened, and houses one of the most impressive natural history displays in North America, go see it!).
Since I was working with the UMNH, I got valuable input from several of the researchers who worked on the specimens. They were able to provide additional information - I won't go into the nitty-gritty of it (although you may if you would like), but I wanted to point out that the end result was quite a surprise to me. And little is more satisfying than when you are really surprised at the end of a skeletal reconstruction.

Resulting skeletal in hand, you can compare it to the most recent studies of the therizinosaur family tree, as well as the excellent research being done by Lindsay Zanno and Peter Makovicky on the origin of plant-eating in theropod dinosaurs, and Falcarius starts to tell an interesting tail about the order in which therizinosaur traits appeared.
Falcarius appears to already be specialized for browsing for high forage. Given the lack of an enlarged gut for fermentation it probably preferred to seek out higher-quality plant matter, like fruiting bodies or seeds. The partially upright stance appears concurrently with a widening of the passage through the pelvis (not visible in side view) allowing move guts into that area, causing the center of gravity to sit further back despite the elongation of the neck.
The large hand claws (from which the authors derived the name "sickle-cutter") may have allowed Falcarius to pick up small prey, but they also may have served as defense for a fairly slow animal with small teeth. The first toe is low and long enough to start interacting with the ground, perhaps to provide balance and stability when browsing high. All of these features would be carried to extremes in advanced therizinosaurs, but they seem to already be playing the same (albeit incipient) functional roles in Falcarius.
So with Falcarius we have an animal that at first glance appears inexplicably strange, but when viewed through the lens of where it was coming from (long-bodied small-headed meat eaters) and where it ends up (the upright, pot-bellied therizinosaurs) the combination of traits start to make a lot of sense.
Isn't science grand?
References:
Kirkland, J. I., Zanno, L. E., Sampson, S. D., Clark, J. M. & DeBlieux, D. D., 2005. A primitive therizinosauroid dinosaur from the Early Cretaceous of Utah. Nature, v435, pp 84-87.
Zanno, L. E. 2006. The pectoral girdle and forelimb of the primitive therizinosauroid Falcarius utahensis (Theropoda, Maniraptora): Analyzing evolutionary trends withing Therizinosauroidea. Journal of Vertebrate Paleontology, v26 n3, pp 636-650.
Zanno, L. E. 2010. Osteology of Falcarius utahensis (Dinosauria: Theropoda): characterizing the anatomy of basal therizinosaurs. Zoological Journal of the Linnean Society. v158, pp 196-230.
Zanno, L. E. & Makovicky, P. J., 2011. Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution. Proceedings of the National Academy of Sciences. v108 m1, pp 232-237.