
Happy Holidays!
/
The science and art of paleontology
A blog on dinosaur anatomy and reconstructing the appearance of prehistoric animals.

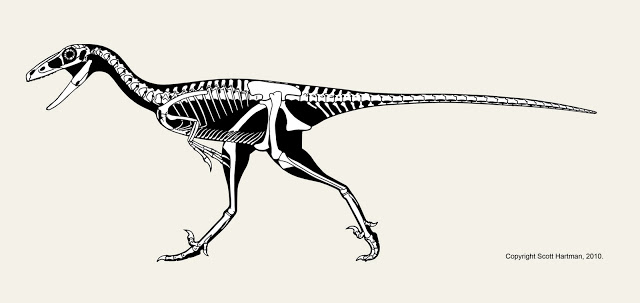
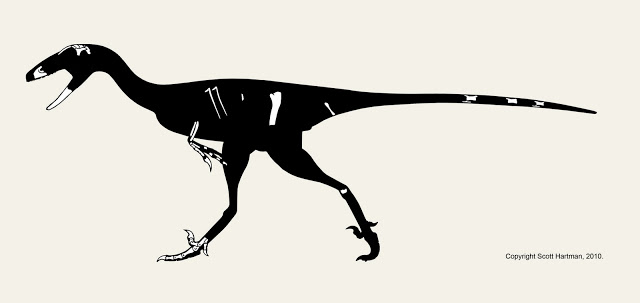
The truly strange looking animal above is Falcarius utahensis. It's an early, omnivorous member of the theropod clade known as therizinosaurs. Not only does it look weird, it's also a bit different from other skeletals you may have seen on the web. Join me after the break for a bit of a discussion about Falcarius, and the challenges I faced with this reconstruction.
I should warn you, this won't be a tutorial on how I make my skeletal reconstructions. That would certainly be a fun series, but it would require quite a lot of time to do properly, so for now it'll have to wait. But there are still several points worthy of discussion.
First off, the animal was discovered in a bone bed of disarticulated individuals. The good news is that most of the individual elements are known, but the down side is the bones aren't all from the same sized animals. That means that cross-scaling is needed to restore the skeleton, but even that presents a challenge; the usual method of cross-scaling involves double-checking the results against the proportions of close relatives. Alas, in this case the fossil record for the base of the therizinosaur family tree isn't well known, and what is known makes it clear that Falcarius has very different proportions than it's closest known relative: Beipiaosaurus.

(Image copyright Greg Paul)
When the original description of Falcarius was published in 2005, it came with the skeletal drawing at left. Obviously I don't agree with those proportions now, but at the time it had been done when fewer bones had been excavated, prepared, and described in detail, so Greg Paul had to try and scale them based on a smaller amount of material to compare with.
In fact, given the difficulty of restoring the proportions I intentionally avoided doing a Falcarius skeletal reconstruction for several years. I might have avoided it all together, but towards the end of my tenure at the WDC we mounted a cast of Falcarius that Gaston Design produced. Working on that skeleton I was able to not only measure and photograph all of the elements, but spend time looking at how the individual elements were matched up. Some parts of the cast's vertebral column are from different sized individuals (an unavoidable consequence of trying to piece together a skeleton from several different individuals). In other cases, vertebrae I had assumed to be from different sized animals were in fact crushed.
.jpg)
In addition to the hands-on data, Lindsay Zanno had been hard at work publishing more detailed information on Falcarius (this is actually notable, as not all researchers are as timely with getting more detailed descriptions of a new animal into print).
As the information piled up I felt that a skeletal was possible to be done. I still didn't tackle it though, as there were plenty of "low-hanging fruit" skeletals that could be done from less-challenging animals.
As luck would have it, I ended up being asked to produce a skeletal of Falcarius for a display in the new Utah Museum of Natural History building (side note: the new UMNH building just opened, and houses one of the most impressive natural history displays in North America, go see it!).
Since I was working with the UMNH, I got valuable input from several of the researchers who worked on the specimens. They were able to provide additional information - I won't go into the nitty-gritty of it (although you may if you would like), but I wanted to point out that the end result was quite a surprise to me. And little is more satisfying than when you are really surprised at the end of a skeletal reconstruction.

Resulting skeletal in hand, you can compare it to the most recent studies of the therizinosaur family tree, as well as the excellent research being done by Lindsay Zanno and Peter Makovicky on the origin of plant-eating in theropod dinosaurs, and Falcarius starts to tell an interesting tail about the order in which therizinosaur traits appeared.
Falcarius appears to already be specialized for browsing for high forage. Given the lack of an enlarged gut for fermentation it probably preferred to seek out higher-quality plant matter, like fruiting bodies or seeds. The partially upright stance appears concurrently with a widening of the passage through the pelvis (not visible in side view) allowing move guts into that area, causing the center of gravity to sit further back despite the elongation of the neck.
The large hand claws (from which the authors derived the name "sickle-cutter") may have allowed Falcarius to pick up small prey, but they also may have served as defense for a fairly slow animal with small teeth. The first toe is low and long enough to start interacting with the ground, perhaps to provide balance and stability when browsing high. All of these features would be carried to extremes in advanced therizinosaurs, but they seem to already be playing the same (albeit incipient) functional roles in Falcarius.
So with Falcarius we have an animal that at first glance appears inexplicably strange, but when viewed through the lens of where it was coming from (long-bodied small-headed meat eaters) and where it ends up (the upright, pot-bellied therizinosaurs) the combination of traits start to make a lot of sense.
Isn't science grand?
References:
Kirkland, J. I., Zanno, L. E., Sampson, S. D., Clark, J. M. & DeBlieux, D. D., 2005. A primitive therizinosauroid dinosaur from the Early Cretaceous of Utah. Nature, v435, pp 84-87.
Zanno, L. E. 2006. The pectoral girdle and forelimb of the primitive therizinosauroid Falcarius utahensis (Theropoda, Maniraptora): Analyzing evolutionary trends withing Therizinosauroidea. Journal of Vertebrate Paleontology, v26 n3, pp 636-650.
Zanno, L. E. 2010. Osteology of Falcarius utahensis (Dinosauria: Theropoda): characterizing the anatomy of basal therizinosaurs. Zoological Journal of the Linnean Society. v158, pp 196-230.
Zanno, L. E. & Makovicky, P. J., 2011. Herbivorous ecomorphology and specialization patterns in theropod dinosaur evolution. Proceedings of the National Academy of Sciences. v108 m1, pp 232-237.

In 1999 Jeff Wilson and Matt Carrano published an excellent paper addressing the phenomena of "wide-gauge" sauropod trackways. For years researchers had been working to explain why sauropod trackways seemed to come in two very different flavors - some of them were very closely spaced...so much so that they would actually overlap on the midline of the track. Other sauropod tracks seemed to show animals walking with their feet spread much further apart.
What were paleontologists to make of this?
One explanation was that the trackways were made by the same type of sauropods that were engaging in different behaviors. In other words, perhaps sometimes a sauropod would walk with its legs close in, while at other times it would use a wide-gauge stance.
Wilson & Carrano proposed that instead the trackways were made by sauropods with different skeletal adaptations. They mustered quite a few lines of evidence, but perhaps the best was that there was a group of sauropods - titanosaurs - that in fact had a much wider pelvis than other sauropods. The paper created a framework for later workers to use when attempting to correlate track makers with fossilized trackways, and is generally a towering success.
But I did want to take issue with one figure of the paper - one that pops up repeatedly at SVP. It is figure 5, demonstrating their interpretation of hing leg stance:

That's Camarasaurus on the left and Opisthocoelicaudia on the right. The clever reader may have already surmised from the title of this post that I think the animal on the right has its legs spread too far apart. But I have a larger issue: both animals have their legs spread much too far apart.
Remember that narrow-gauged trackways actually have their feet fall so close together that they frequently overlap along the midline. There's no way even sauropod "A" could make those tracks in the stance as figured. And this is why I'm bringing this up, because animals generally don't walk around with their legs acting as perfectly vertical beams. If you spend time watching large animals walk away from you, you'd see something like this:

(Elephant image from here, rhino image from here.)
People also move like this, with our vertical limbs generally sloping in toward the midline when we walk or run. There are probably several reasons for this (including mechanical efficiency) but for our purposes here let's just setting on the fact that it happens. Large, straight-limbed graviportal animals tend to walk with the limbs angled inward, not down (and certainly not angled out).
And the trackways also demonstrate this. If you place place sauropods over the actual trackways in question, you end up with a stance more like this:
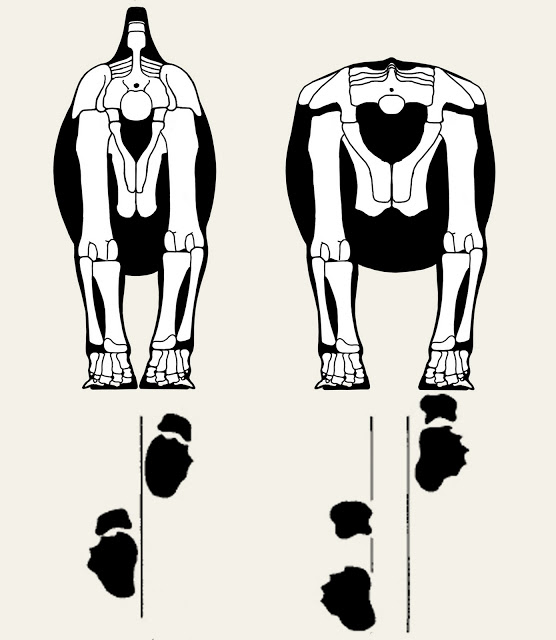
In this case I've put a diplodocid (Supersaurus) on the left, while the animal on the right is scaled to the pelvic dimensions of Opisthocoelicaudia as seen in the original paper. Both animals have the hind legs mostly vertical but gently sloping inward.
This is not to say that sauropods never adopted a pose with their legs spread out a bit; Wilson & Carrano point out that titanosaurs have adaptations that may have allowed them to evert their hind limbs more effectively. They suggest that titanosaurs may have done so when rearing up, or during other activities that require greater stability.
I don't take issue with that, and those sorts of differences in the legs and pelvis may make it possible to tease out further behavioral differences between sauropod groups. But when walking around in their day to day lives both the footprints and modern analogs make a strong case that the limbs should be vertical, and if anything sloping in towards the midline rather than spread away from it.
Reference:
Wilson, J. A, & Carrano, M. T. 1999. Titanosaurs and the origin of "wide-gauge" trackways: a biomechanical and systematic perspective on sauropod locomotion. Paleobiology, 25(2), pp. 252-267.

Ok, time for a quick anatomy lesson: Despite what you may have heard, bird knees do not bend backward. Nor, in fact, do the knees of any tetrapod perform this trick. Given the role of the knee in locomotion, it's not even clear how such a reversal could evolve after the initial "knee bend" direction was settled upon several hundred million years ago.
Why bring the anatomical equivalent of a fairy tale? Well, it's a fairly common misconception. So common, in fact, that it was recently enshrined by none other than Scientific American. So let's see if we can clear this up with some simple diagrams:

Really, it's not.
It's a surprisingly common mistake. When looking at living birds many people fail to realize that part of the leg is hidden on a bird; the upper leg barely moves, and along with the knee it is actually buried up under the feathers of the wing and body. Birds also have quite long ankles, leaving their ankle joints in roughly the position we'd expect the knees to be on a human. Like this:

The key here is that people are plantigrade animals, while all theropods (including birds) are digitigrade. That means that human ankles are flat on the ground, and in our case our knees are roughly in the middle of our legs. In birds and other digitigrade animals (most dinosaurs and many mammals, like dogs, deer, and horses) it's only the toes that contact the ground. The ankle joint is well up off the ground, and the knee is is actually in the upper 1/3rd of the leg. And in birds the thigh is actually even a smaller part of the leg, and as mentioned above is also mostly hidden under feathers. Here's a look comparing the same leg portions of a human (in two poses), a dog, and an extinct bird:

(Presbyornis copyright Scott Hartman, other skeletals modified after Charles Knight.)
In the diagram the thigh bones (femora) are all colored red, the shin bones and proximal ankle bones are colored blue, the "foot bones" (the distal tarsals and the metatarsals) are green, and the individual toe bones are yellow. Notice that the femur of the extinct bird Presbyornis is small and very high, and the knee (the joint between the red and blue bones) is up where it would be hidden by the body and wings. But please also notice...the knee is bending the same way as ours. And the same way as everything else that has a spine and walks on land.
Which brings us back to the article posted by Scientific American. I don't want to go too far with "gotcha" blogging, but Scientific American is generally one of the more highly regarded popularizers of science on the web and in print. I took a look the other pieces published by the author, and she seems like a solid reporter that just happens to have made a mistake. Journalists don't get science degrees (and even if they did that would only be one subject), so this should not be construed as an attack.
But at the same time, this is a serious error. It's like reporting that September has 32 days in it, or that the Red Sox clinched a playoff spot this week. Only worse, as it's perpetuating a myth that gets passed around as "common knowledge". I attempted to bring this to the attention of the relevant parties shortly after it was reported, both on Google+, where SciAm blog editor Bora Zivkovic has been making effective use of the new social network, and the author's Twitter account (which is frequently used).
Despite near real-time feedback, days have gone by with no correction. Making a mistake is understandable, but failing to correct it is not. Scientific American has written hundreds of articles on the state of science education, and has often been an effective advocate for ways to improve it. But the authority they derive comes from their attention to scientific detail, so I hope we will now see a quick correction without further delay.
It's my pleasure to introduce the newest member of the troodontid family:Talos sampsoni. Named for paleontologist Scott Samspon, Talos was described by Lindsay Zanno and others in the wonderful open source journal Plos ONE. Talos is the first troodontid to be named from the Kaiparowits Formation of Utah, making it about 76 million years old. Cross-sections of the long bones suggest that the animal was between four and six years old, and while it hadn't stopped growing, it appeared to be reaching reproductive age at a smaller size than it's close relative Troodon.
Of note is that the specimen had a bone in its second toe that was injured and partially healed. Since it appears to have been injured from violent trauma, it's consistent with with the idea that the "switchblade" toe was used in a way that could result in such an injury (presumably either attack or defense). Also, since the rest of the foot shows no indication of the sort of limping or other adjustments you see from a prolonged foot injury, it also reinforces the idea that troodontids walked with the second toe off the ground (as shown above).
I was producing the above skeletal reconstruction for the Utah Museum of Natural History, who has constructed a new building filled with exciting displays that will opened Fall 2012. It should be a state of the art exhibit that everyone should go see if they get the chance. Dr. Zanno was working on describing Talos at the time, so I worked with her to incorporate the data into the skeletal. There is quite a bit of troodontid material from the Kaiparowits, but only a fraction of it can be confidently assigned toTalos at this time. Restricting ourselves to the type specimen looks something like this:

You could be forgiven if your first reaction is "that's not very complete!". But I was thrilled. Why? It's hard to remember sometimes, since Troodon is illustrated frequently, but the published material that Troodon is usually based on is really scrappy. Here is a quick and dirty example of what was published in Dale Russell's 1969 paper:
To create the reconstruction (then called Stenonychosaurus) ofTroodon
that inspired him to create his infamous dinosauroid sculpture, Dale Russell had to combine all of the material from the Asian taxa Saurornithoides (as seen below) just to create a usable composite:

That made the size of the head and pelvis more clear, and provided more back and tail vertebrae to scale from, but look at how "dotted" the outlines of the limbs are - there still wasn't a good guide for scaling the limbs of any of these advanced troodontids. A few unpublished specimens are out there, but none of those provided definitive limb proportions. Talos finally does that.
So in addition to the interesting conclusions on biogeography, ontogeny, and mode of life, Talos should be exciting to illustrators because it provides the first definitive glimpse of the limb proportions of advanced troodontids. They aren't radically different from the proportions Dale Russell speculated on in the 1960s (although the forelimbs are shorter than some people have speculated since then), but the good news is that we now know for sure. In fact at this point the only serious unknown is the neck, although there are plenty of more basal troodonts to base that on.

If you watched Episode II of Dinosaur Revolution, you may have laughed at this Ornitholetes, who I'll refer to as Ichabod. This may seem like an odd scene to pick for a scientific discussion, but I think it actually has something useful to teach. Also, I'm partially responsible. I should be clear, the story idea was not mine (that's above my pay grade), but it's something that was run by me, and I did not try to shoot it down (and still wouldn't).
Why? I think it's reasonable.
No wait, let me explain: I'll grant you that there isn't much in the professional literature on the subject on the subject of "headless running" in animals, but from some criticisms I've read I think people maybe thinking about this the wrong way (i.e., wondering about the distribution of "headless-running" in birds like the behavior it's an advanced condition).
Vertebrates as a group have one of the more centralized nervous systems among animals (with some arthropods and especially some cephalopods as the other contestants in the "flexibility over redundancy sweepstakes"), but tetrapod nervous system evolution in general is a story of progressive centralization that (so far) culminates in mammals. Even in humans, with our gobs of ridiculously calorie-hungry centralized gray matter, we still have autonomous reactions that don't require the brain to be involved (as anyone knows who's burned themselves and jerked their hand away before they felt the pain). That said, we have gone a long way down the path of nervous system centralization, and if you cut a mammal's head off you may get some twitching but it won't run around; our limbs literally cannot coordinate themselves without the brain's involvement (although morbidly it does appear that the head itself retains some coordination afterwards, if medieval reports are true that heads react for up to a 15 seconds after a beheading).
This degree of centralization is the derived condition, not the primitive one. So it seems unlikely that chickens are special here, except in as much as they more frequently get clean beheadings in the presence of human observers than most other birds (a quick Google search shows that turkey's exhibit this as well). This should be true of lizards, crocs, etc too (diapsids as a whole). So with enough experimental trials I'd fully expect an Ornnitholestes to do a good headless chicken impression. Now, I'll grant you that this would require a pretty clean bite on the allosaur's part, and the odds of observing it in the wild would not be very high. But the sequence was devised as be a surprising bit of humor in a scenario that was possible, not probable.
Given those parameters it seems reasonable enough to me.

Paleontologist and paleoartist Scott Hartman provides a wealth of anatomical data and skeletal drawings of dinosaurs, and information on how prehistoric animals looked in life.















{kind=link}