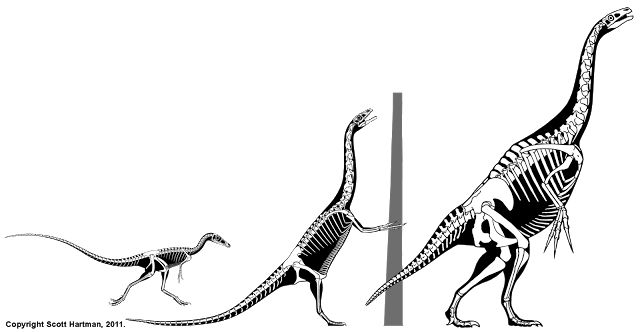
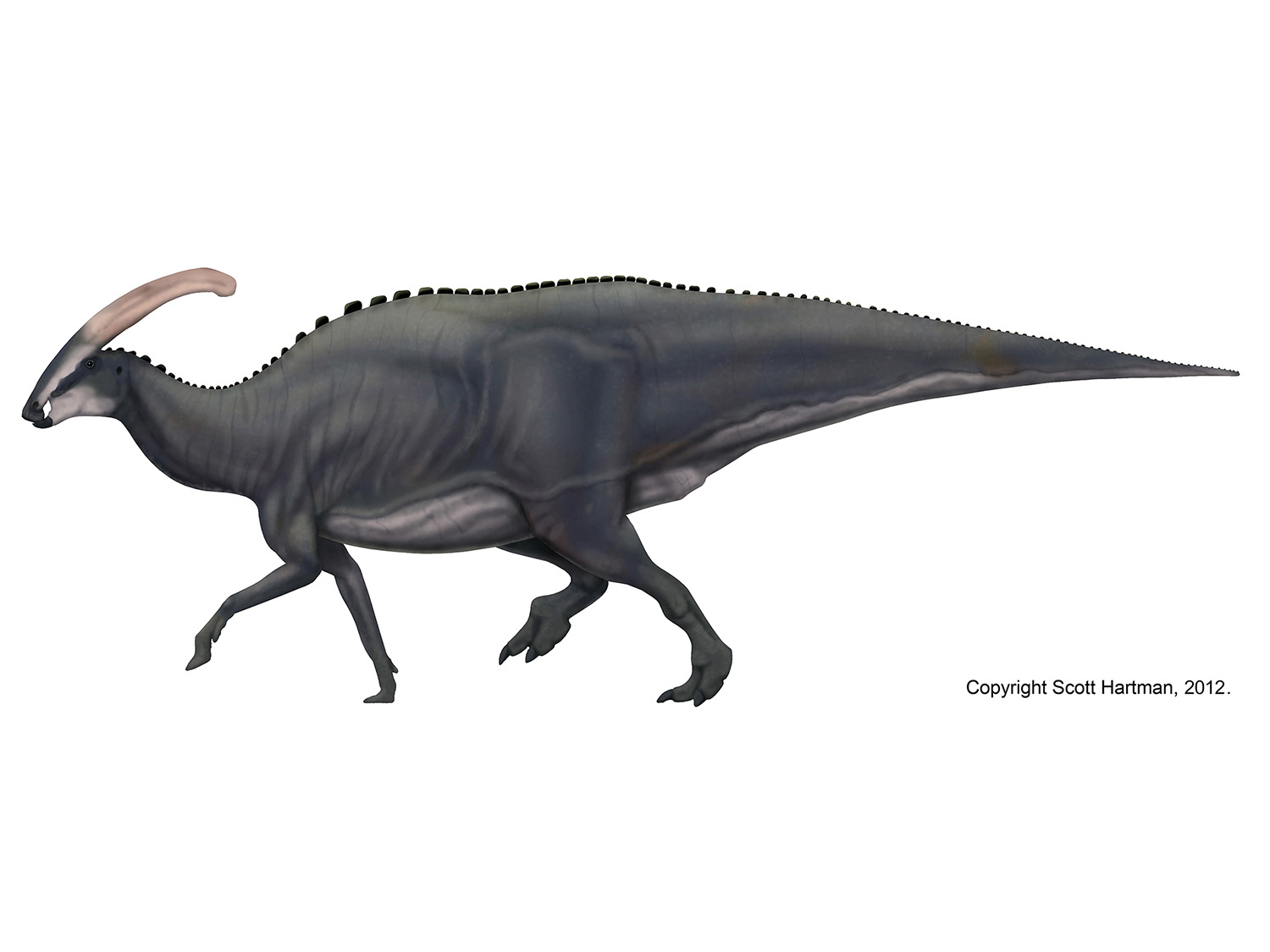
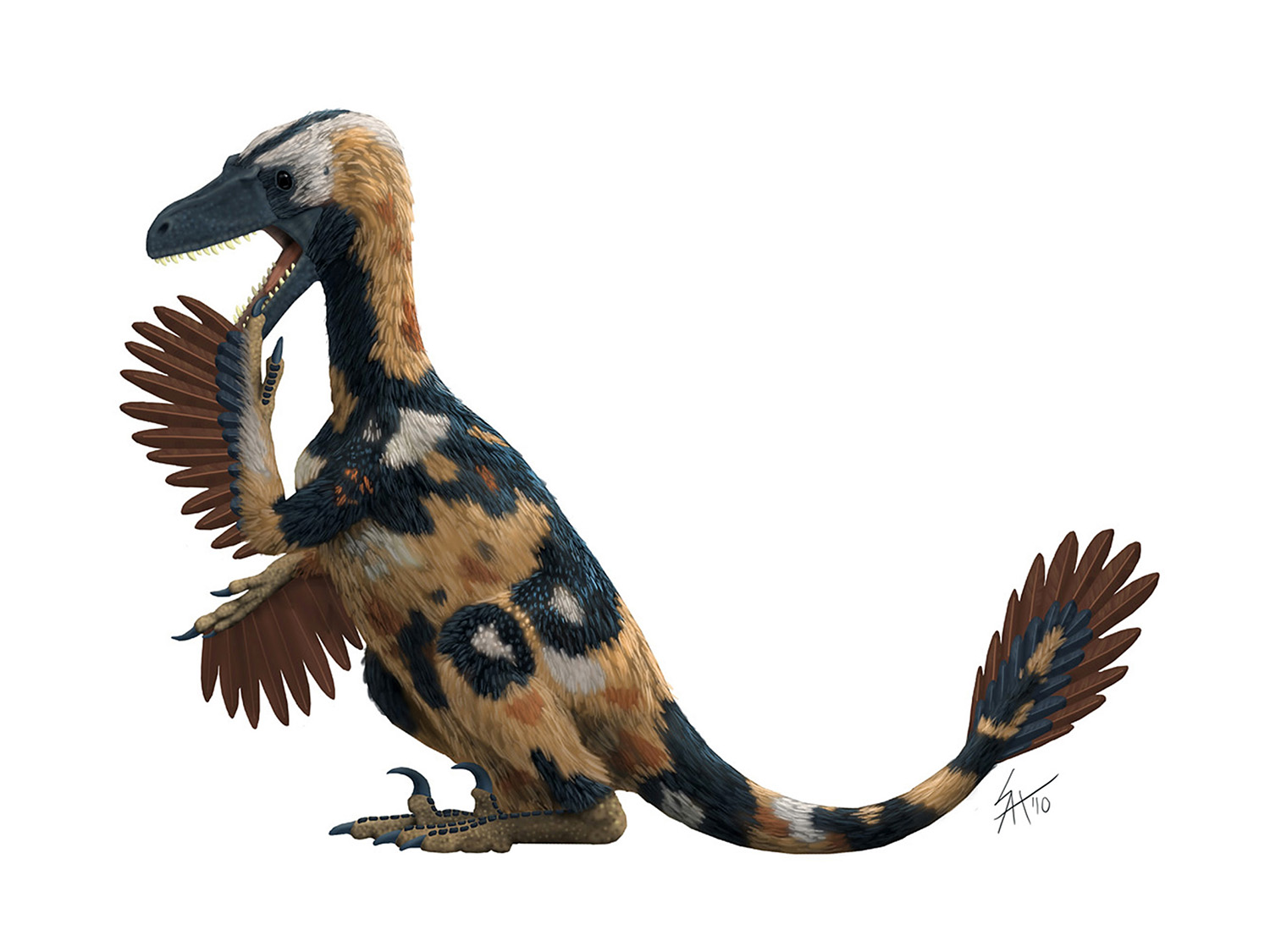Well, it's 2012, so the Great Skeletal Repose of 2011 must officially come to an end. Most of the bipedal skeletals in my collection have been reposed much like this Velociraptor. I had plenty of things to say, and we saw some great discussion by guest writers, but when it comes right down to it, the issue feels incomplete without some sort of summary as to how I got here, and what's left to do. So let's take a quick look at where things stand now...
To some degree, the walking pose shown above was selected by acclimation. When I started to show off different types of poses, by far the most popular was the walking pose you see above. And I want this pose to be one that other scientific illustrators can feel free to adopt, so widespread can only enhance that proposition. But it wasn't just a popularity contest - there are several practical reasons why this walking pose became the winner:
The Good
1) Utility - Greg Paul of course will continue to use his pose, and a number of previously published skeletals by other authors (including all of mine up until last year) had adopted the same pose. By selecting the walking pose the retracted left leg remains unchanged, allowing for a easy comparison of the proportions. This was probably the biggest factor.
2) Functional Aesthetics - The old pose of animals sprinting along at a lively clip tended to impose a specific hypothesis of activity on the animals. While in some cases dashing along may have been quite likely, it still required that a research who wanted to use the skeletals to swallow that hypothesis, whether they agreed with it or not. With Velociraptor that wasn't unlikely, but with larger theropods it became needlessly controversial, and with graviportal taxa like therizinosaurs the results could be laughable.
3) Laziness - I prefer "efficient" over "lazy", but no matter what label you place on it, this pose required a minimal amount of alteration to my existing skeletals. And the stark reality is that when you have to repose hundreds of technical illustrations that time adds up. Quickly. Of course this reason would not have been sufficient if it weren't for the more important points 1 & 2, but it sure was a nice bonus.
Does that mean I'm thrilled with the outcome?
Yes and no. Since I'm starting with a blank slate, it would have been fun to come up with something truly distinctive. Yet the allure of the new wasn't worth sacrificing how useful the skeletals were for comparative purposes. Also, thanks to lots (and lots) of time spent pondering the issue, and to the extra stimulus provided by Mike Habib and Jon Conway's excellent guest posts, I've come to some unsettling questions that are still unresolved.
The Bad
1) Whither goest utility? - As I wrote way back in the halcyon days of...spring 2011, after I announced that I would relinquish the old pose and select a new one I got many requests that I refrain from this. Almost all of them were concerned with losing the ability to easily contrast my skeletals to others after the repose. I was (and am) sympathetic to that plight, and selected a pose that minimized the "damage".
Yet I also had a disconcerting realization: Both Greg Paul and myself had been varying the pose of the forelimbs for years and no one ever bothered to complain about the loss of utility. Sometimes we've both illustrated maniraptorans with the arms flexed into their folded-wing pose, sometimes not. Less advanced theropods obviously never adopted such a pose, so their arms often hang listlessly. My tyrannosaurs went from a similar "hanging out" pose to one that reflected the work done by Lipkin and Carpenter. Why was the issue never broached? Are forelimbs less important for comparative purposes than hind limbs? Or do we naturally gravitate towards the larger structure because of how our visual cortex's work? I don't know, but I'm unsatisfied by the discrepancy.
2) Is variety the spice of life? - Jon Conway also brought up a good point in his guest article, that there isn't a single pose that best serves every need. This is certainly true, and while I still feel that within groups making poses similar is useful, I also have to admit that in some ways the job has just begun.
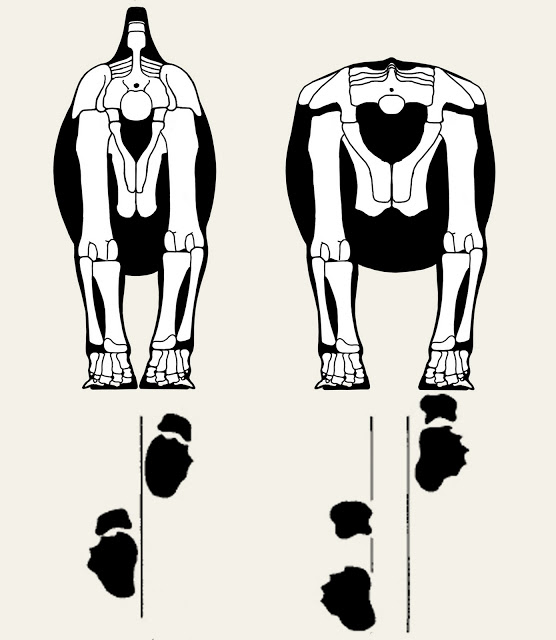
3) That job has just begun - Oh yeah, and another thing. Turns out there are quadrupedal dinosaurs too. Who'd have thunk it, eh? Despite the obvious and objective superiority of theropods, prosauropods, and basal ornithiscians, there's still a lot of four-footed critters in my skeletal collection, and I'm going to have to come up with a pose for them as well. Two actually, since the graviportal species will need a pose that is different from the quadrupeds with flexed limbs. Ah well, that just means that 2012 will also need to have a Great Skeletal Repose as well.
An end and a beginning
So we've come full circle. I've adopted a pose for the bipedal dinosaurs, but still have to come up with (two!) new poses for quadrupeds. I still am very much interested in soliciting outside opinions on the subject, but I also want the blog to move back to posts about anatomy and reconstruction, rather than a continuing series of posts on the technical issues behind selecting a pose. So expect to see the occasional progress report on the quadrupeds, but don't expect it to dominate space on the blog this year.
If you have a strong opinion on the subject, don't hesitate to email me (or use that Gchat thing). In the mean time, I really do have a series of upcoming posts on Acrocanthosaurus and Spinosaurus anatomy, as well as the trials and tribulations of reconstructing skeletals in multiple views.
Stick around, won't you? 2012 should be an interesting year.











.jpg)